 药学进展
药学进展延伸阅读:行业动态 | 先声药业皮下注射PD-L1抗体Envafolimab临床试验数据将在ASCO大会发布
2020年5月15日,先声药业宣布,其与江苏康宁杰瑞生物制药有限公司(以下简称“江苏康宁杰瑞”)、思路迪(北京)医药科技有限公司(以下简称“思路迪医药”)达成战略合作的新一代PD-L1抗体KN035(通用名:Envafolimab)“单药在中国高微卫星不稳定性(MSI-H)...全文>>
专家介绍:燕茹
博士,澳门大学中华医药研究院暨中药质量研究国家重点实验室副教授,博士生导师。2008年加入澳门大学,成立澳门首个药代动力学实验室。主要研究方向为中药代谢和动力学,致力于建立符合中药临床用药和整体作用特点的中药药代、药效和毒性的研究方法和基于体内过程的中药质量评价体系。目前研究重点为肠道菌群代谢及肠道菌-宿主共代谢为基础的疾病治疗靶标发现、中药干预评估及创新中药研发。主持国家自然科学基金、澳门科技发展基金、广东省自然科学基金及澳门大学科研基金等课题十余项。在Analytical Chemistry、Drug Metabolism and Disposition、Applied Environmental Microbiology、Journal of Ethnopharmacology等SCI期刊上发表研究论文80余篇。担任中国药理学会药代专委会青委会副主任委员、分析药理专委会委员及药学会医药生物分析专委会委员, Journal of Ethnopharmacology杂志编委。获2016中国中西医结合学会科学技术奖一等奖(排名第6)和2018澳门科技发展基金技术发明奖三等奖(排名第3)。
个人网页:https://sklqrcm.um.edu.mo/ruyan/.
正文
肠道菌群代谢药物研究进展
[摘要]长期以来,由于肠道菌群本身的复杂性以及培养方法和分析技术的限制,肠道菌群在药物代谢和体内处置中的作用未得到应有的重视。近年来,随着技术的快速发展,围绕肠道菌群的大量研究将对肠道菌代谢和肠道菌-宿主共代谢的认识提高到前所未有的深度。肠道菌群不仅能够直接代谢许多药物,还通过宿主、菌群、药物之间复杂多维的相互作用间接改变药物代谢,从而影响个体对药物治疗的响应(效应、毒性、耐药性等)。肠道菌群结构及代谢功能受多种因素的影响。综述近期肠道菌群代谢药物研究领域的重要进展及研究趋势,以期推进精准治疗,促进药物发现及新的治疗策略的出现。
人体表面及内部栖居着种类众多、数量巨大的微生物,其中95%以上生活在胃肠道内,其数量和组成因栖居的微环境而异,沿消化道逐渐增多,在结肠达到最高(1010~1011CFU·mL-1)。成年人的肠道菌群组成以厚壁菌门(Firmicutes)、放线菌门(Actinobacteria)和拟杆菌门(Bacteroidetes)为主。健康人的肠道菌群组成尽管存在较大个体差异,但构成其核心菌群的14个属均相同。肠道菌群失调与多种疾病息息相关。
肠道菌具有强大的代谢功能,由肠道微生物基因编码的酶的代谢活性,远远超过人体基因组编码的蛋白的代谢活性。肠道菌除了可直接分解利用营养物质,维持肠道内环境稳态及身体代谢平衡,还通过肠道菌代谢或肠道菌-宿主共代谢参与药物的处置,影响药效或毒性。宏基因组学技术揭示了肠道菌群的多样性,但也发现仍有约80%的人肠道微生物是未知的,这严重阻碍了对肠道菌功能的研究;绝大部分口服药物主要在小肠上段吸收,因此,传统的采用粪便菌的体外代谢研究难以准确评估肠道菌对药物代谢的贡献;此外,肠道菌群结构和功能受多种因素影响,宿主、菌群和药物之间也存在复杂的相互作用。然而药物研发阶段的药物代谢和动力学(DMPK)研究中往往忽略以上因素,导致肠道菌对药物代谢的贡献被低估。目前,有明确报道的肠道菌群参与代谢的药物仅有50多个,但最近一项研究系统评估了76个代表性菌株对271种口服药物化学修饰的能力,发现2/3被测药物经与至少一种菌株孵育后药物水平显著降低,单一菌株能够代谢11~95种药物,表明肠道菌群对口服药物代谢的影响更为广泛。随着肠道菌群研究的持续升温,人们对肠道菌群的代谢能力以及宿主-菌群-药物互作的认识不断深入,推动了肠道菌代谢药物研究的迅速发展。本文综述肠道菌群代谢药物研究领域的最新进展,并总结研究趋势及其对药物研发和治疗的影响。
1肠道菌代谢药物的反应类型
还原和水解是肠道菌催化的最主要反应类型,其结果是分别抵消了宿主代谢催化的以促进药物消除为目的的氧化和结合反应。此外,肠道菌群还能催化官能团转移、裂解反应等(见表1)。
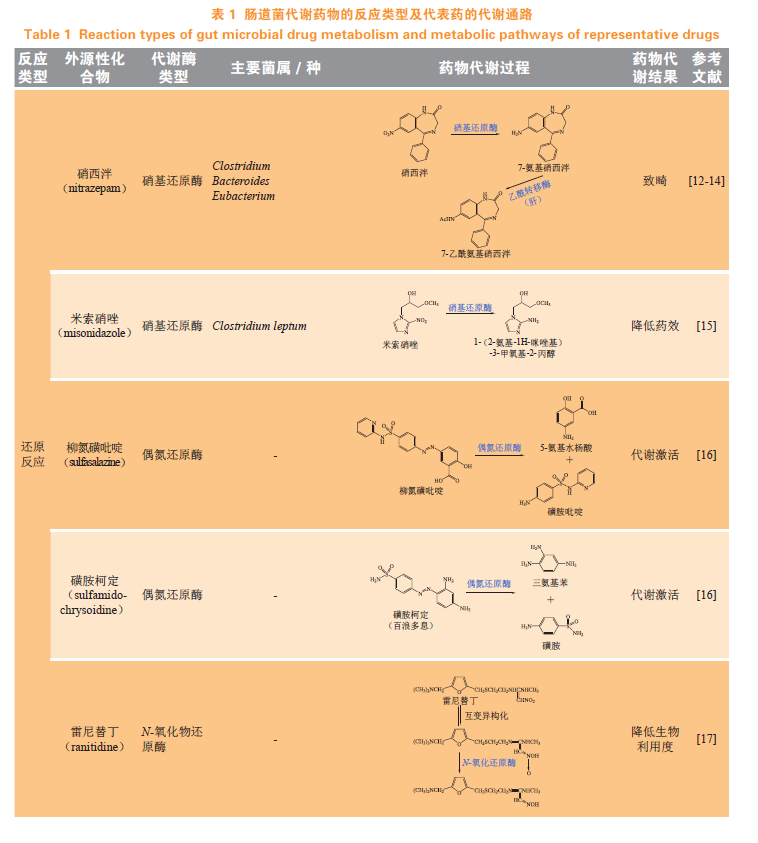
1.1还原反应
肠道菌还原酶能够利用各种辅基、辅酶[烟酰胺腺嘌呤二核苷酸(NADH)、烟酰胺腺嘌呤二核苷酸磷酸(NADPH)、黄素单核苷酸(FMN)等]来介导电子转移,通过降低药物极性、改变电荷及亲电性来影响药物的活性及在体内的保留时间。人肠道菌还原酶催化的硝基和偶氮化合物的还原反应较为常见。此外,肠道菌还可还原亚砜基、α,β-不饱和羧酸等官能团。
1.1.1硝基还原硝基是一种良好的药效官能团,常见于抗肿瘤药物中,具有很强的亲电子能力,在体内易被酶催化还原,发挥药理作用或产生毒性。硝基还原酶主要有2种类型:Ⅰ型硝基还原酶可从NADH或NADPH转移2个电子至底物中的硝基,将其转变成氨基;Ⅱ型硝基还原酶可将单个电子转移至硝基形成硝基阴离子自由基。Bacillus spp.、Clostridium spp.、Escherichia coli等菌能编码表达硝基还原酶的基因。苯二氮䓬类药物硝西泮(nitrazepam)经口给予怀孕大鼠后,在肠道内可被硝基还原酶还原为7-氨基硝西泮,后者在肝脏中代谢生成7-乙酰氨基硝西泮。Clostridium、Bacteroides、Eubacterium属的菌均具有还原硝西泮的酶活性,其中,从人体肠道内分离出的Clostridium leptum能够显著催化硝西泮的硝基还原反应。此外,Haemophilus influenzae表达的硝基还原酶能不同程度地还原硝基化合物,如氯霉素(chloramphenicol),可将其结构中的硝基还原成氨基。米索硝唑(misonidazole)与大鼠肠道菌混合物培养或与Clostridium perfringens单独培养,能被还原成一种氨基衍生物。
1.1.2偶氮还原偶氮基(-N=N-)常用于前药的化学结构修饰,可被还原生成活性代谢物。肠道中部分好氧菌及部分厌氧菌,如Escherichia coli、Enterococcus faecalis、Staphylococcus aureus、Bacillus cereus、Clostridium perfringens等均能表达偶氮还原酶。尽管不同菌中偶氮还原酶的结构和功能活性存在差异,但均能还原裂解偶氮键。美沙拉嗪(mesalazine)是一种氨基水杨酸类抗炎药,能直接作用于肠道,用于治疗炎症性肠病,但其口服后在小肠几乎完全吸收,经肝脏代谢失活,无法到达结肠发挥疗效,因此,基于5-氨基水杨酸(5-ASA)设计出活性较低且不易口服吸收的偶氮类前药柳氮磺吡啶(sulfasalazine)、奥沙拉嗪(olsalazine)和巴柳氮(balsalazide)。这类药物主要在结肠处被代谢,释放出活性代谢物5-ASA,达到靶向治疗溃疡性结肠炎的效果。此外,磺胺类抗菌药磺胺柯定(sulfamidochrysoidine),即百浪多息(商品名),也是一种前药,其结构中的偶氮基经偶氮还原酶裂解,生成具有抗菌活性的磺胺(sulfanilamide)。
1.1.3N-氧化物还原肠道菌介导的N-氧化物还原影响宿主对人类免疫缺陷病毒抑制剂BILR355的代谢。早期研究认为BILR355主要由细胞色素P450酶系(CYPs)3A亚家族(CYP3A)酶代谢,利托那韦(ritonavir)与BILR355联用后通过抑制CYP3A酶使BILR355的体内暴露增加、疗效增强;但同时在血浆中检测到一种新的主要代谢物BILR516,实验发现该产物是经肠道菌还原BILR355结构中的N-氧化物生成中间产物BILR402,后者被宿主醛氧化酶进一步代谢生成BILR516。雷尼替丁(ranitidine)和尼扎替丁(nizatidine)均属于组胺类H2受体拮抗剂,用于治疗消化性溃疡等,肠道菌可通过还原其结构中的氧化胺,降低药物吸收与生物利用度。前药氧洛哌丁胺(loperamideoxide)同样可经该代谢途径生成活性化合物洛哌丁胺,发挥止泻作用。
1.1.4亚砜还原亚砜基(>S=O)也是一种在药物结构中常见的基团。环氧化酶(COX)抑制剂磺吡酮(sulfnpyrazone)和非甾体抗炎药舒林酸(sulindac)结构中的亚砜基均可被人或兔的粪菌还原生成硫化物,其中人肠道菌对舒林酸的还原能力更强,生成甲硫基化合物发挥药效;抗生素可抑制肠道菌对亚砜基的还原作用。体外研究发现,奥美拉唑(omeprazole)的亚砜基也可被大鼠肠道菌还原,但由于奥美拉唑口服后在小肠吸收良好,很难到达消化道远端,因此难以评估体内肠道菌对该药代谢的贡献。
1.1.5腙还原腙是含有R1R2C=NNH2结构的有机化合物。人口服艾曲波帕(eltrombopag)后可在尿液中检测到腙裂解代谢物。当该药与啮齿动物的盲肠内容物或人粪便样品在厌氧条件下孵育时可检测到还原产物,而与CYPs、肾或肝微粒体等孵育则未检测到该代谢物。
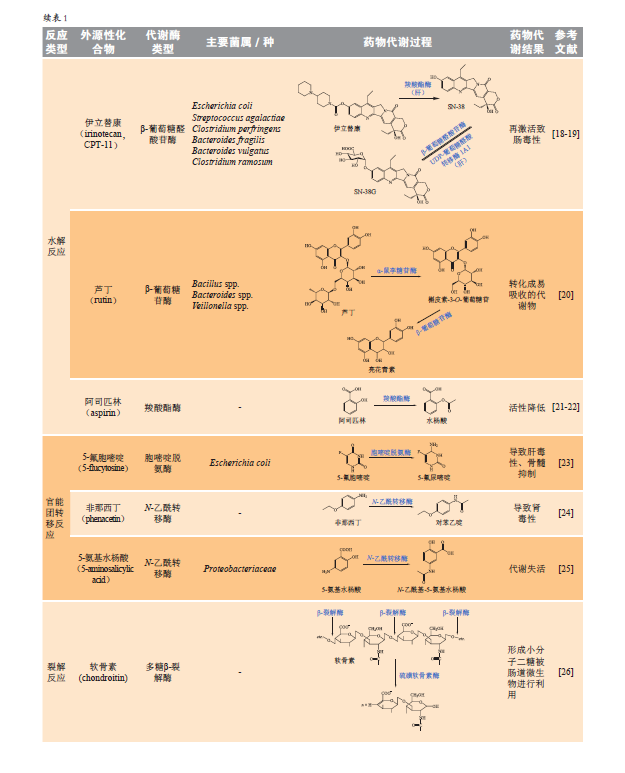
1.2水解反应
肠道菌主要表达β-葡萄糖醛酸苷酶(GUS)、β-葡萄糖苷酶、羧酸酯酶(CES)等多种代谢酶催化水解反应,改变药物及其代谢物的理化性质与活性。
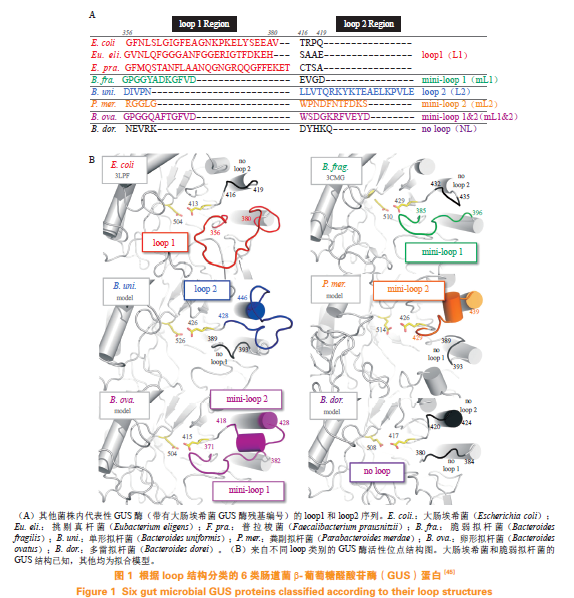
1.2.1β-葡萄糖醛酸苷酶GUS在肠道菌中分布广泛,主要存在于厚壁菌门、变形菌门、疣微菌门和拟杆菌门中。目前,从人类微生物组计划GI数据库中鉴定出共3013个、279种独特的菌群基因编码的GUS蛋白。通过对肠道菌GUS蛋白的序列进行比对,发现其结构中预测α/β水解酶折叠的活性位点和核心区域序列高度保守,而临近活性位点的loop结构序列为菌GUS蛋白独有,存在高度可变性,根据loop结构可以将肠道菌GUS蛋白大致分为6类(见图1)。目前,已知loop结构的差异与GUS底物选择性存在一定联系。通常具有loop1结构的GUS对小分子葡萄糖醛酸苷底物(如4-硝基苯基-β-D-葡萄糖醛酸、双氯芬酸葡萄糖醛酸)的催化能力更强,对GUS选择性抑制剂UNC10201652的敏感程度也更高;而loop结构更开放的GUS(如mL1、L2、mL2以及NL类GUS)可以代谢大分子葡萄糖醛酸苷如乙酰肝素;此外,NL类GUS能特异性水解N链葡萄糖醛酸底物瑞格非尼葡萄糖醛酸苷(regorafenib-glucuronide)。结肠癌的一线治疗药物伊立替康(irinotecan)经静脉注射后可在肝脏被羧基脂酶水解,生成活性代谢物SN-38,后者在肝脏尿苷二磷酸-葡萄糖醛酸转移酶1A1(UGT1A1)作用下生成葡萄糖醛酸苷结合物SN-38G,但该结合物经胆汁排泄进入肠道,被肠道菌GUS水解,生成SN-38,导致肠道黏膜受损引发腹泻。研究显示,双氯芬酸、吲哚美辛、布洛芬等多个非甾体类抗炎药通过相似机制引起肠道损伤。
1.2.2β-葡萄糖苷酶人体自身存在多种β-葡萄糖苷酶,包括溶酶体β-葡萄糖苷酶、小肠黏膜刷状缘的乳糖酶-根皮苷水解酶(LPH)和胞质β-葡萄糖苷酶(CBG)。肠道菌群中多数革兰阳性厚壁菌门也具有β-葡萄糖苷酶活性,尤其是梭菌类群ⅩⅣa的Eubacterium rectale和Roseburia spp.,以及梭菌类群Ⅳ的Faecalibacterium prausnitzii。该类酶可水解天然产物中广泛存在的葡萄糖苷类结合物,形成更易吸收和/或有生物活性的苷元。黄酮类单糖苷可经小肠LPH或CBG酶以及肠道菌β-葡萄糖苷酶共同催化去糖基化产生更易吸收的苷元。但对于非单糖取代的槲皮素类糖苷(如芦丁和金丝桃苷),肠道自身的LPH和CBG均无法分解,只有通过肠道菌的糖苷酶水解产生苷元。中药黄芪主要皂苷成分黄芪甲苷经肠道菌水解产生具有端粒酶激活作用的环黄芪醇,而其主要黄酮成分毛蕊异黄酮7-氧-β-D-葡萄糖苷经肠道菌代谢后主要以苷元形式吸收,后者经肝UGT1A9催化生成的3'-葡萄糖醛酸苷可表现出更强的促血管新生作用。
1.2.3羧酸酯酶CES能够催化酯、酰胺、氨基甲酸酯或硫酯键断裂。CES在体内分布广泛,在肠、肝和肠道菌中均有表达。构成宿主CES蛋白结构中活性空腔内的基本催化三联体为丝氨酸(Ser)、组氨酸(His)和谷氨酸(Glu),而菌CES的催化结构是由Ser、His和天冬氨酸(Asp)组成。宿主CES被认为是经典的异源化合物代谢酶,可水解许多具有酯基的药物;而菌CES作用于相对分子质量小的水溶性酯,在疏水化合物(如胆固醇)的内源性加工过程中也起着重要作用。Kim等发现阿司匹林可被肠道菌CES水解生成水杨酸,与抗菌药氨苄西林联用能抑制肠道菌CES活性,增强其抗血栓作用。
1.3官能团转移反应
肠道菌代谢酶通过亲核取代,可催化官能团在底物之间转移以改变化合物的极性,影响药物的吸收、排泄、毒性和药理活性。
辛伐他汀(simvastatin)是一种内酯类前药,通过抑制羟甲基戊二酸单酰辅酶A(HMG-CoA)来降低胆固醇水平,其与人结肠微生物体外培养可发生去甲基化反应。在人粪便中能检测到肠道菌代谢福他替尼(fostamatinib)后的O-去甲基化和二羟基化产物。抗真菌药物5-氟胞嘧啶可经肠道菌中的胞嘧啶脱氨酶转化生成5-氟尿嘧啶。口服非那西丁(phenacetin)除了经肝脏CYPs(主要为CYP1A2)催化O-脱乙基生成对乙酰氨基酚(acetaminophen)发挥药效外,还能被肠道菌乙酰转移酶代谢,生成具有肾毒性的对苯乙啶。地尔硫䓬(diltiazem)结构中的乙酰酯基可被肠道菌Bacteroidesthe taiotaomicron编码的酶(BT_4096)代谢生成去乙酰化产物。此外,5-ASA经肠道菌催化乙酰化,导致抗炎作用显著减弱。
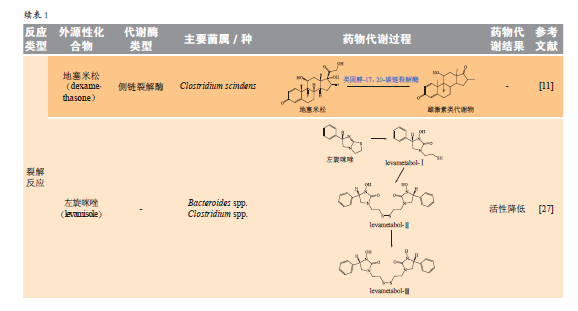
1.4裂解反应
肠道菌可表达裂解酶来直接裂解C-X键(X为C、O、N、S、P或卤素),释放可利用的小分子代谢物或降低药物的生物利用度。
1.4.1β-裂解人体肠道微生物能够编码超过5000种多糖裂解酶(PLs)。PLs能够通过和CES共同作用,裂解多糖中位于羧基β位的糖苷键,从而获得小分子单糖。此外,肠道微生物还具有C-S键β-裂解酶。一些食物、药物等外源性物质经CYP450酶代谢后形成具有潜在毒性的代谢产物,部分活性代谢中间产物在肝中与谷胱甘肽形成结合物,该结合物经胆汁排泄进入肠道,被肠道菌群的C-S键β-裂解酶催化裂解。例如,对乙酰氨基酚在肝内生成的谷胱甘肽结合物经胆汁排泄进入肠道,经肠道菌β-裂解酶代谢形成巯基取代产物,该产物可被重吸收进入肝,被肝线粒体甲基转移酶甲基化,生成易蓄积的甲硫基加合物。
1.4.2侧链裂解侧链裂解主要发生在固醇类药物。地塞米松(dexamethasone)、氢化泼尼松(prednisolone)、可的松(cortisone)等药物的侧链碳链可被Clostridiums cindens表达的碳链裂解酶裂解生成雄激素形式的代谢产物。萘丁美酮(nabumetone)属于非固醇类抗炎药,口服后结构中的侧链被肠道菌裂解生成活性产物6-甲氧基-2-萘基乙酸(6-MNA)。
1.4.3开环氧化左旋咪唑(levamisole)与肠道菌体外培养生成多种噻唑环开环的氧化产物,使其药理活性减弱。甲硝唑(metronidazole)由肠道菌Clostridium perfringens代谢开环生成N-(2-羟乙基)-乙酰胺酸和乙酰胺。抗惊厥药唑尼沙胺(zonisamide)结构中的苯并异唑环可被Clostridium sporogenes开环成2-氨磺酰基乙酰苯酚(SMAP)。
1.5其他
CYPs是一类单加氧酶,其催化的I相反应是药物在体内代谢转化的关键步骤。CYPs主要在哺乳动物的肝、肠道黏膜中表达。此外,在多个细菌中也有CYPs表达,具有非常强的代谢活性。哺乳动物的CYPs是一类膜结合型蛋白,催化代谢反应需要氧的参与,而菌表达的CYPs是可溶性蛋白,催化过程不需要氧参与。目前,对菌CYPs代谢药物的研究报道较少。肠道菌Streptomyces spp.表达的CYPs能代谢非甾体类抗炎药氟比洛芬(flurbiprofen)生成羟基化产物,除和宿主CYPs一样催化4位羟基化外,还能催化3位的羟基化或甲氧基化。
2肠道菌代谢药物的结果
2.1代谢激活(前药)
许多药物由于难于吸收、代谢不稳定、具有刺激性或毒性,常被修饰成无/低活性的前药来提高生物利用度、避免毒副作用。其中一些前药能被肠道菌代谢激活(activation),释放出活性代谢物,从而发挥药效,如5-ASA的前药(奥沙拉嗪、巴柳氮、柳氮磺吡啶)和磺胺类化合物的前药百浪多息经肠道菌代谢分别释放出5-ASA和磺胺。洛伐他汀(lovastatin)的γ-内酯环结构水解产生的2-羟基洛伐他汀酸是其治疗高胆固醇血症的活性成分。Yoo等观察到在抗生素处理后大鼠的血浆中2-羟基洛伐他汀酸降低,推断是肠道菌介导了γ-内酯环的水解活化。然而,最近研究显示,洛伐他汀γ-内酯环结构的水解是一个pH依赖的自发的转化过程,而肠道菌并不能催化γ-内酯环水解,而是介导了2-羟基洛伐他汀酸的分解,使药效降低。
肠道菌能够通过代谢药物,影响化疗药的治疗效果。对30个化疗药物的体外分析显示,大肠埃希菌显著增强6个药物(替加氟、磷酸氟达拉滨、5-氟胞嘧啶、6-巯基嘌呤-2'-脱氧核糖苷、AQ4N和CB1954)的体外杀伤肿瘤细胞的活性,同时分别检测到大肠埃希菌水解氟达拉滨的核苷酸糖苷键以及还原CB1954(羟胺及胺衍生物)的相应代谢物,而瘤内注射大肠埃希菌则显著提升了CB1954治疗小鼠的中位生存率。
植物药中广泛存在的糖苷类成分(皂苷、黄酮糖苷、蒽醌类糖苷、二苯乙烯苷等)由于极性较强,不易被吸收。肠道菌表达的葡萄糖苷酶或GUS可水解一些糖苷类化合物的糖苷键,生成疏水性较强的苷元,促进吸收,或产生活性代谢物。例如人参皂苷类口服生物活性很低,主要通过肠道菌(如双歧杆菌、拟杆菌、梭菌等)β-葡萄糖苷酶催化脱糖,生成有同样活性或活性更强的次生代谢物,如原人参二醇型人参皂苷的主要代谢物CompoundK具有更强的抗肿瘤、抗炎等作用,是口服人参皂苷后血中检测到的主要药物相关成分。黄芩苷是葡萄糖醛酸苷,口服后可在肠道中由肠道菌GUS水解为易被小肠上皮细胞吸收的黄芩素,后经肠/肝UGT酶重新转化为黄芩苷进入血液循环,发挥其药理作用。蒽醌类化合物(如大黄素、大黄酚、大黄素甲醚等)是大黄中一类主要的生物活性成分,天然存在形式主要是葡萄糖苷,经口服后在体内主要以葡萄糖醛酸苷形式存在。研究显示,在右旋葡聚糖硫酸钠引起的大鼠慢性结肠炎中,由于肠道菌GUS活性降低,肠肝循环被抑制,导致大黄蒽醌成分结合物的系统暴露降低。
2.2代谢再激活
肠道菌代谢导致的药物再激活[即代谢再激活(reactivation)]普遍存在于进行肠肝循环的药物以及许多天然产物成分。宿主肠/肝代谢酶催化药物发生结合反应,其代谢物(葡萄糖醛酸苷)可通过胆汁排泄进入肠道,被肠道菌GUS水解释放出药物,部分经肠道重吸收。因此,宿主肠/肝UGT-肠道菌GUS代谢轴协同作用决定了这些药物的体内处置过程及药效发挥。部分药物(如吗啡)的原型系统暴露增加可提高药效,或活性代谢物(如抗癌药伊立替康)在肠局部的暴露增加产生毒性。此外,用作预防肾脏移植后排斥反应的免疫抑制剂霉酚酸酯(MMF)作为霉酚酸(mycophenolic acid,MPA)的前药,经肝脏CES1和CES2代谢为MPA后,经肝UGT酶-肠道菌GUS作用,可增加活性代谢物MPA的系统暴露。然而上述天然糖苷类成分经肠道菌水解以苷元形式吸收后,后者经肠/肝代谢酶催化,进入肠肝循环过程,促进苷元形式的利用。
2.3代谢失活
强心苷类药物地高辛(digoxin)通过结合人心肌细胞中的Na+/K+ATP酶来治疗充血性心力衰竭等心脏疾病。肠道菌Eggerthella lenta可使其代谢失活(detoxifcation)生成二氢地高辛,测序分析发现一种只存在于Eggerthella lenta中的强心苷还原酶操纵子,能编码2种强心苷还原酶(Cgr1和Cgr2),催化还原地高辛结构中的α,β-不饱和内酯,从而减弱药物与Na+/K+ATP酶的结合能力,降低疗效。左旋多巴(levodopa)是治疗帕金森病的药物,在肠道被Enterococcus faecalis中的酪氨酸脱羧酶(TyrDC)代谢生成强极性的多巴胺,后者进一步被Eggerthella lenta中多巴胺脱羟基酶(Dadh)代谢生成3-酪胺,两者均无法透过血脑屏障,使药效降低。
2.4代谢致毒
硝西泮经肠道菌硝基还原酶和肝N-乙酰转移酶代谢生成的7-乙酰氨基硝西泮具有致畸性。非那西丁经肠道菌脱乙酰酶基生成的对苯乙啶与长期服用非那西丁导致的肾炎有关。5-氟尿嘧啶可以直接被肠道菌中广泛分布的脲嘧啶磷酸核糖转移酶代谢,生成5-氟尿苷单磷酸(5-fluorouridine monophosphate),破坏RNA。京尼平苷(geniposide)可被肠道菌β-葡萄糖苷酶水解,生成具有肝细胞毒性的对苯二酚。
2.5药物耐受
化疗药物的耐药一直是阻碍癌症治疗的一个难题,最近研究发现肠道菌代谢能导致部分化疗药物耐药。核苷酸类似物吉西他滨(gemcitabine)用于治疗胰腺癌、肺癌、乳腺癌或膀胱癌,其与大肠埃希菌共孵产生乙酰化代谢物,瘤内注射大肠埃希菌显著降低吉西他滨治疗组小鼠的生存率。结肠癌小鼠模型瘤内γ-变形杆菌表达的长型胞苷脱氨酶(CDDL)使吉西他滨脱氨失活;另外,在高达76%胰腺管型腺瘤患者的肿瘤组织样品中检测到菌DNA,其中半数以上的片段来自γ-变形杆菌中的肠杆菌科和假单胞菌科,而绝大多数患者新鲜肿瘤组织中的菌能使人结肠癌细胞株RKO和HCT116对吉西他滨产生完全耐受。
2.6药物-药物相互作用
目前,对肠道菌代谢药物引起的药物-药物相互作用(drug-drug interactions,DDI)尚缺乏系统深入的研究。肠道菌嘧啶核苷磷酸化酶在Bacteroides spp.中活性较高,能水解抗病毒药物索立夫定(sorivudine)生成溴苯尿嘧啶(bromovinyluracil),后者透过肠道进入肝后与二氢嘧啶脱氢酶结合,造成对该酶的不可逆抑制,降低其对联用的5-氟尿嘧啶的降解,导致5-氟尿嘧啶蓄积产生毒性。
3影响肠道菌代谢药物的因素
肠道菌基因组较人体基因组更易受其他因素的影响而发生结构和多样性的改变,进而影响代谢功能。影响肠道菌组成的因素众多,如围产期因素(出生及喂养方式、遗传、肠黏蛋白糖基化等)、饮食营养、疾病、生活方式(抽烟、饮酒、熬夜等)、用药(尤其是抗生素)、地理、环境等。多种疾病包括代谢性疾病(如糖尿病)、肝病(如脂肪肝)以及炎症等普遍存在菌群多样性降低的问题,低纤维西式饮食也往往导致菌群多样性降低甚至某些重要的菌消失。抗生素抗药性是当代医学一个日益严重的现象,已引起人们的广泛关注。最近针对市场上1000种不同类别的非抗生素类药物的体外研究发现,约24%的受试药物(包括临床常用的抗精神性药物、质子泵抑制剂、激素及抗肿瘤药物等)在临床治疗浓度能够对以肠道共生菌为主的40个代表菌株中的至少一个产生明显的生长抑制,这与临床观察到的这些药物的副作用相一致。对来自欧洲不同区域的2个健康成年人群的粪便样品组成及变异因素分析显示,用药对菌群结构变异的贡献最大。由此可以推测,许多非抗生素类药物也可能对菌群结构产生影响,促进某些耐药性的产生,并影响药物代谢、药效和药物毒性。
对肠道菌的生理、病理作用重要性的认识推动了靶向菌群的各种干预措施的出现,包括通过抗生素联用、益生菌、益生元、粪菌移植等直接影响菌群结构和多样性以及靶向干预特定肠道菌代谢通路。例如,胆碱利用基因簇(choline utilization cluster,cut)存在于多种肠道菌中并能编码一对微生物蛋白CutC和CutD(CutC/D)来介导三甲胺(TMA)生成,服用CutC/D的共价抑制剂碘甲基胆碱可显著降低小鼠血浆氧化三甲胺(TMAO)的水平,改善胆碱饮食诱导形成的强化血栓等。然而,这些干预措施本身也可能改变肠道菌的组成和功能,影响菌群对药物的代谢。如经口给予小鼠益生菌Lactobacillus reuteriK8,检测到Bifdobacterium和Clostridium数目显著增加,伴随降解对乙酰氨基酚的硫酸酯酶(sulfatase)、芳基硫酸盐磺基转移酶(arylsulfate sulfotransferase)活性显著升高和GUS活性下降,经口给药对乙酰氨基酚的系统暴露水平相应降低。给予实验性结肠炎模型大鼠人参多糖能重建菌群平衡,部分逆转肠道菌β-葡萄糖苷酶活性的降低,增强人参皂苷Rb1的经口吸收和肠道菌代谢,从而增强了Rb1及其次生代谢物的系统暴露。
与宿主代谢相似,肠道菌结构和代谢随饮食习惯和进食时间表现出一定的生物节律性,引起菌群代谢的周期性变化,反过来影响宿主代谢的生物节律。因此,旅行中的时差可以打破肠道菌的生物节律引起菌群扰动。小鼠肠道黏膜相关菌会随着生物周期而变化,导致其黑暗期的数量是光照期的10倍。菌群的昼夜节律性变化甚至会对药物的副作用产生影响,夜间给予小鼠对乙酰氨基酚会引发更严重的肝损伤,原因在于肠道菌在夜间分泌更高水平的1-苯基-1,2-丙二酮,使肝脏GSH含量减少,提高了肝的易感性。
此外,给药途径、制剂技术(控释技术、靶向给药等)、肠肝循环等可以通过影响药物在肠道的驻留和与菌群的接触,改变菌群代谢药物的程度和生物利用度。
目前,尚不清楚这些因素导致的肠道菌群组成变化的信息与临床观察到的药物反应(代谢、效应和毒性)的变动之间是怎样的关系,认识到许多干预措施本身也可能改变肠道菌的组成和功能,则使这一问题变得更为复杂。然而,肠道菌群本身的功能冗余性(redundancy)和多效性(pleiotropy),又能使菌群维持一定范围内结构变化的弹性及功能的韧性。因此,哪些影响因素能最终在菌群功能上留下印迹,引起行为表型和生理特性的变化,将成为肠道菌群的稳态调节研究的新方向。
延伸阅读:行业动态 | 先声药业恩度®(重组人血管内皮抑制素)二项研究入选2020ASCO年会壁报展示
受COVID-19疫情影响,全球最大的肿瘤临床研究年会ASCO(American Society of Clinical Oncology)今年将于美国时间5月29日-5月31日举行网络会议。在ASCO近日公布入选的研究摘要中,来自先声药业的恩度®(重组人血管内皮抑制素)二项研究入选2020ASCO年会壁报...全文>>
4靶向肠道菌代谢药物的治疗策略及药物研发
肠道菌群如何影响药物代谢、药效或毒性并对其进行量化已成为目前研究的热点领域之一。因此,精确编辑肠道菌群,调节其代谢,将促进新的治疗策略的出现,推动药物研发及精准治疗。目前,靶向肠道菌代谢药物的治疗策略及药物研发主要有以下几个方面:
1)抗生素联用策略降低耐药性或毒性:针对瘤内菌引起的吉西他滨耐受,同时给荷瘤小鼠腹腔注射抗生素环丙沙星,能恢复吉西他滨的抗肿瘤活性,表明菌代谢介导的药物耐受可以通过同时给予抗生素的方式改善。同样,怀孕大鼠经口给药硫酸新霉素、盐酸四环素和杆菌肽,在不影响肝硝基还原酶的前提下,几乎完全抑制了盲肠内容物中硝基还原酶活性,可显著降低硝西泮的致畸性。
2)抑制肠道菌药物代谢酶以提高药物系统暴露或减毒:选择性抑制细菌GUS可以有效减少伊立替康肝代谢物SN-38G在肠道中的再活化,降低其毒性产物SN-38在肠道中蓄积,从而缓解伊立替康的肠毒性。采用相同策略也能使非甾体类抗炎药对小鼠胃肠道的损伤减轻。这些研究发现推动了对肠道菌GUS的分布、结构、功能和抑制特性的系统表征,并促进了抑制剂的研发。针对肠道菌降解肽类药物的问题,Sasaki等利用柠檬酸作为细菌降解阿奇替林(azetirelin,促甲状腺激素释放激素类似物,一种神经保护药物)的抑制剂,与制剂增强剂正月桂基-β-D-麦芽吡喃糖苷以及阿奇替林一起制备成肠溶胶囊,使阿奇替林在犬中的生物利用度从15%提升至高于40%。左旋多巴需进入大脑转化为多巴胺才能发挥对帕金森病的治疗作用,针对肠道Enterococcus faecalis中的TyrDC代谢导致左旋多巴脑中暴露降低的问题,设计出TyrDC脱羧反应的强效抑制剂(S)-α-氟甲基酪氨酸(L-酪氨酸类似物),其能显著增加Enterococcus faecalisMMH594定植的悉生小鼠血中左旋多巴的浓度。
3)利用肠道菌药物代谢设计新的药物或递送系统:对于一些生物特性尤其是溶解性较差的药物,可以将药物以磷酸酯或硫酸酯前药的形式给药,如泻药硫酸钠(sodium picosulfate)就以二硫酸盐的形式使用,依赖肠道菌的磺基转移酶将其转化为4,4'-二羟基二苯基-(2-吡啶基)-甲烷发挥作用。针对柳氮磺吡啶经肠道菌水解产生具有副作用的磺胺吡啶,开发出一种大分子药物递送系统,该系统通过间隔基团将药物5-ASA在5位偶氮连接至聚乙烯胺,产生水溶性的偶氮聚合物链接药物,利用结肠病变部位肠道菌催化还原代谢,释放5-ASA,改善该药的治疗指数。此外,采用基于多糖的结肠靶向递送系统可实现益生菌的定点释放,也为设计结肠靶向药物提供了新思路。
5肠道菌群代谢药物研究的趋势及展望
综观近期肠道菌群代谢药物的研究呈现以下趋势:
1)研究系统化:多学科手段、系统性研究。由于目前肠道菌多数不能通过体外培养获知其代谢功能,研究通常利用对个别已知菌的目标代谢功能分析,结合功能性宏基因组学以及生物信息学,获取肠道菌代谢药物的基因/酶分布信息,用以指导针对微生物组的合理治疗干预措施。例如,为鉴定肠道菌中催化L-多巴脱羧反应的酶,MainiRekdal等首先提出L-多巴脱羧反应需要磷酸吡哆醛依赖性酶的假说,并在肠道菌基因组中搜索候选基因,从粪肠球菌中鉴定出保守的TyrDC,进一步通过遗传和生化实验证实,TyrDC可同时使其优选底物酪氨酸及L-多巴脱羧;在此基础上,发现酪氨酸的模拟物(S)-α-氟甲基酪氨酸能够抑制TyrDC、粪肠球菌以及帕金森病患者的肠道菌群样品的L-多巴脱羧作用。此外,结合多种预测模型和计算工具,可评估胃肠道不同位置微生物对药物的影响。Zimmermann等利用无菌小鼠、药物代谢酶缺失细菌构建限菌小鼠模型,定量测定宿主和肠道菌群对药物代谢的贡献,成功检测了抗病毒药索利夫定在不同部位的代谢情况,发现核苷类抗病毒药溴夫定转化为肝毒性代谢物的过程71%由肠道菌群完成,进一步结合药代动力学模型实现对药物/代谢物在不同个体系统暴露的准确预测。
2)研究精细化:具体到菌株/酶/基因水平。多数情况下,介导药物代谢的菌和酶并不清楚。但多学科技术手段的应用及培养组学的出现,促进了肠道菌代谢药物的机制研究。如鉴定出肠道菌代谢L-多巴的路径,包括经粪肠球菌的TyrDC转化为多巴胺,然后经迟缓埃格特菌的DadhR506变异,代谢为m-酪胺;吉西他滨的耐药性是由于肿瘤内的γ变形菌门表达的CDDL代谢使药物失活引起;而拟杆菌属中普遍存在的由bt4554基因编码的嘌呤核苷磷酸化酶介导了溴夫定的代谢。这些机制的深入研究促进了靶向肠道菌药物代谢环节的药物发现及新的药物联用策略。
3)纯培养和培养组学(culturomics)迅速发展:由于仍有80%以上的人肠道微生物是未知的,单独依靠宏基因组技术无法阐明微生物的代谢功能,近期纯培养技术重新获得重视。采用多重培养条件结合基质辅助激光解吸/电离飞行时间型质谱仪(MALDI-TOF)及16SrRNA测序等技术鉴定微生物的培养组学快速发展,使得可培养的人体细菌增加了几百种,推动了对人体肠道细菌的营养偏好、代谢特性研究以及对肠道菌代谢口服药物的系统性研究。
4)药物微生物组学的出现:药物微生物组学是研究菌群与药物反应/代谢互作的新兴领域。宿主、菌群、药物之间存在复杂多维的互作关系,药物可影响肠道菌群的组成和功能,肠道菌群除了直接代谢药物,还通过影响代谢、免疫、炎症等,间接影响药物反应和生物利用度,而遗传和饮食等因素均影响肠道菌群和药物代谢。通过无菌动物、微生物定植或粪菌移植研究证实,肠道菌能通过释放可溶性因子、细菌外膜囊泡(OMVs)或代谢产物影响宿主代谢酶表达或活性。色氨酸的肠道菌代谢产物吲哚丙酸通过TLR4信号通路作用于调控药物代谢和转运的重要感受器孕烷X受体(pregnane X receptor,PXR)。肠道菌群能调控肝肠P-糖蛋白等转运子的表达,从而影响药物肠吸收或在消除器官的暴露。此外,菌群代谢物还可以通过竞争机制影响宿主药物代谢。如对甲酚(p-cresol)能够通过竞争磺酸化酶,降低扑热息痛的磺酸化代谢,使体内产生较高对甲酚的患者发生肝毒性的风险增大。另一方面,菌群是机体免疫平衡和炎症反应的重要调节者,炎症/感染通过细胞因子介导机制使宿主代谢能力严重受损。因此,药物微生物组学研究为临床重新解读药物的系统暴露行为提供了更宏大深入的机制背景,也为应用生理药动学模型(PBPK)预测药物吸收、分布、代谢和排泄(ADME)带来更为复杂的考量,可促进个性化治疗。
肠道菌代谢药物的研究仍面临巨大挑战。肠道菌群在肠道内的分布存在着空间和组成的不均一性。不同肠段的pH值、氧分压及肠上皮细胞生理状态等存在差异,比如小肠段的pH值较结肠低(pH6~7vspH7)而氧分压较结肠高(pO233mmHgvs<33mmHg),导致小肠部位定植菌的核心类群不同,多样性低。对于大部分主要在小肠吸收的药物,通过粪菌研究获得的药物代谢结果无法反映小肠部位菌群代谢药物的真实情况。但是目前较难获得小肠部位的菌群样品。多数体外方法过于简化,无法反映肠道生态系统的特性,如特定微生态环境、pH梯度的差异、动态的微生物-组织互作。人体肠道微生物生态模拟系统可模拟完整的胃肠道动态环境,通过精细调控系统的环境参数(pH、氧分压、滞留时间等)获得复杂、稳定的微生物群落,成为研究药物代谢、食品发酵、肠道菌代谢的重要工具。微生物成像技术通过和其他方法相结合(如生物正交化学荧光标记、质谱成像等)来动态监测宿主菌群变化、更准确地鉴定相关代谢物及这些代谢物的空间分布,进一步推动了这一领域的发展。最近受到较多关注的胃肠道类器官、器官芯片微生理系统能够较好地模拟体内系统,提高研究效率,可以用于肠道菌代谢和宿主-微生物互作研究。可消化的电子胶囊可提供胃肠道氧、氢、二氧化碳的实时信息,用作研究肠道菌代谢的化学生物感应器,指示局部发酵情况。
对肠道菌群的代谢能力、肠道菌-药物-宿主互作关系的认识不断深入,将大力推进以下几个方面的研究:1)药物处置的菌群-宿主的合作、协调反馈机制;2)肠道菌组成及功能个体差异大并受多种因素影响,将肠道菌群代谢纳入PBPK模型,实现对个体药物代谢及系统暴露的差异更为精准的预测,Zimmermann等的工作为定量评估肠道菌代谢药物及药代动力学预测做了很好的示范;3)培养组学技术结合功能基因组、生物信息学在菌/酶/基因水平阐明药物代谢机制;4)体外培养条件下单一菌的研究很难复制胃肠道复杂环境下的代谢能力,人肠道微生态模拟技术将获得更大发展;5)肠道菌纯培养、蛋白质重组、结合分子模拟等技术系统研究肠道菌代谢酶的代谢、诱导或抑制等特性。对肠道菌群参与药物代谢的分子机制的理解,将最终推动靶向菌群代谢的药物研发和个性化治疗新策略的出现和完善。
综上所述,对肠道菌群的代谢功能以及宿主、菌群、药物之间复杂多维互作关系的认识不断深入,使我们能够重新审视并认识肠道菌在药物的代谢和体内处置以及药效或毒性中扮演的重要角色。功能性宏基因组学、培养组学、微生物成像、微生态模拟、分子模拟以及生物信息学等多种技术手段的发展及综合应用,推动肠道菌代谢药物研究向着系统性、精细化快速发展。阐明肠道菌群影响药物代谢的分子机制并对其进行量化,将肠道菌群代谢纳入PBPK模型,实现对个体药物代谢及系统暴露的差异更为精准的预测,将促进临床对微生物组的合理干预及精准治疗,推动靶向菌群代谢的药物研发和治疗新策略的出现和完善。
关于药学进展
感谢您阅读《药学进展》微信平台原创好文,也欢迎各位读者转载、引用。本文选自《药学进展》2020年第2期。
《药学进展》杂志是由中国药科大学和中国药学会共同主办、国家教育部主管,月刊,80页,全彩印刷。刊物以反映药学科研领域的新方法、新成果、新进展、新趋势为宗旨,以综述、评述、行业发展报告为特色,以药学学科进展、技术进展、新药研发各环节技术信息为重点,是一本专注于医药科技前沿与产业动态的专业媒体。
《药学进展》注重内容策划、加强组稿约稿、深度挖掘、分析药学信息资源、在药学学科进展、科研思路方法、靶点机制探讨、新药研发报告、临床用药分析、国际医药前沿等方面初具特色;特别是医药信息内容以科学前沿与国家战略需求相合,更加突出前瞻性、权威性、时效性、新颖性、系统性、实战性。根据最新统计数据,刊物篇均下载率连续三年蝉联我国医药期刊榜首,复合影响因子0.760,具有较高的影响力
《药学进展》编委会由国家重大专项化学药总师陈凯先院士担任主编,编委新药研发技术链政府监管部门、高校科研院所、制药企业、临床医院、CRO、由金融资本及知识产权相关机构百余位极具影响力的专家组成。
《药学进展》编辑部官网:www.cpupps.cn;邮箱:yxjz@163.com;电话:025-83271227。欢迎投稿、订阅!
● 想回顾《药学进展》编委会主办和协办过的精彩活动吗?请戳这里!
→
→
→
→
→
→
→
→
→
→
点一下你会更好看耶
延伸阅读:行业动态 | 阿斯利康慢阻肺双联支气管扩张剂百沃平®令畅®在华获批
2020年5月18日,中国上海 ——阿斯利康今天宣布,中国国家药品监督管理局正式批准双联吸入支气管扩张剂百沃平®令畅®[i](通用名:格隆溴铵/福莫特罗吸入气雾剂)用于慢性阻塞性肺疾病(慢阻肺),包括慢性支气管炎和/或肺气肿患者的维持治疗,以缓解症状。 百...全文>>
本文来源:药学进展 作者:药学进展 免责声明:该文章版权归原作者所有,仅代表作者观点,转载目的在于传递更多信息,并不代表“医药行”认同其观点和对其真实性负责。如涉及作品内容、版权和其他问题,请在30日内与我们联系


 我们沟通的很顺畅
我们沟通的很顺畅 电话已拨通,无人接听
电话已拨通,无人接听 这个电话号码是空号
这个电话号码是空号